
傳染性法氏囊的病原學與免疫應答
發布時間:2025-02-18 發布人:華南生物 分類:技術專欄
傳染性法氏囊病是由雙RNA病毒科的禽雙RNA病毒屬的傳染性法氏囊病毒(IBDV)引起的一種高度傳染性的疾病,呈急性并伴有嚴重免疫抑制,主要影響青年雞,造成重大的經濟影響。該病于1957年首次爆發,1962年美國正式報告。早期該病的特點是廣泛性的腎臟損傷,當時被稱為禽腎病。該病造成的巨大經濟損失來源于兩個因素,它通過破壞法氏囊中含有IgM的發育中的B淋巴細胞,造成嚴重的免疫抑制,降低體液免疫應答能力,使雞只更易受到感染,并易導致疫苗接種失敗;3周齡或青年雞,出現60%以上的死亡率。該病潛伏期短,約3-5天,小于3周齡的雞沒有臨床癥狀,但導致免疫抑制。IBDV導致的免疫抑制程度,取決于感染雞的年齡,2周齡的雞比大日齡的雞免疫抑制更嚴重。
一、基因組結構和病原學
傳染性法氏囊病毒的基因組為分段的雙鏈RNA(dsRNA),片段分為A和B,病毒粒子55-60nm,無囊膜,二十面體對稱衣殼。B片段較小,編碼病毒蛋白1(VP1,95kDa)。A片段(2.9kb-3.4kb)具有兩個部分重疊的開放閱讀框(ORF)。其中,較小的ORF編碼非結構蛋白VP5(17kDa),該蛋白與病毒的致病性和病毒在細胞間傳播有關;較大的ORF編碼一個110kDa的多聚蛋白,通過自裂解產生3個多肽:pVP2(48kDa)、VP3(32kDa)、VP4(28kDa)。VP4作為一種具有絲氨酸-賴氨酸蛋白酶活性的蛋白,介導了這一過程。
VP1蛋白是RNA依賴的RNA聚合酶,在病毒粒子中以游離的形式存在,并且該基因組連接蛋白(VPg)附著在兩個基因組片段的正鏈的5’端。研究表明,超強毒vvIBDV致病型的VP1基因序列形成了一個獨特的簇,表明它可能來自于B片段的基因重組。VP1蛋白由于其在病毒復制效率中的作用而調節病毒毒力,通過針對VP1蛋白的RNA(DNA載體為基礎)體外干擾實驗來抑制病毒復制。
外衣殼蛋白VP2是主要結構蛋白,約占病毒總蛋白的51%(Heetal, 2009)。它至少有2個中和表位,可誘導產生中和抗體。它負責抗原變異、適應組織能力和病毒毒力。VP2成熟過程中的2種短肽產物在病毒附著和胞漿內易位期間,決定病毒顆粒在細胞質膜的組裝和破壞過程。蛋白質被折疊成三個關鍵的結構域:基底、外殼和突起。VP2并不是超強毒中唯一的毒力決定因素,VP1蛋白是另一個毒力決定因素(Qi等,2013)。
內衣殼蛋白VP3(32kDa)是Y型三聚體,約占病毒總蛋白的40%,形成病毒蛋白組裝的衣殼內部支架。它具有群特異性和少量的中和抗原表位,與VP1相互作用,通過其羧基末端結構域與病毒基因組物質相互作用。有學者認為,VP3蛋白通過與病毒顆粒的幾乎所有成分(自身、VP2、VP1和兩個基因組dsRNA)的相互作用,促進病毒復制和子代病毒的產生。
VP4(28kDa)是一種病毒自催化蛋白酶,是一種可溶性蛋白,它利用不含ATP酶結構域的絲氨酸-賴氨酸催化二元體,將多聚蛋白裂解為單獨的VP2、VP3和VP4蛋白。VP4在IBDV血清型和毒株中是保守的。該蛋白通過在組裝過程中不斷修飾C端末梢幾個小肽,在pVP2蛋白的成熟過程中發揮重要作用。
病毒非結構蛋白VP5是一種II類膜蛋白,具有細胞質N端和細胞外C端結構域。它在所有的IBDV血清1型株中具有強堿性和半保守性,并富含半胱氨酸。VP5可誘導法氏囊病變,在病毒傳播和釋放中發揮重要作用。該蛋白在細胞膜內積累,導致細胞活力下降。在體外培養中,VP2與VP5可誘導細胞凋亡。
二、抗原漂移和基因組RNA突變
外衣殼蛋白VP2可誘導產生中和抗體。VP2上抗原表位所在的區域具有高核苷酸變異性,表明其易受抗原轉換和漂移的影響。研究表明,超強毒vvIBDV的毒力、組織嗜性和致病表型的決定因素是由VP2蛋白中253、279、284位的一些氨基酸殘基控制的。反向遺傳學研究數據表明,VP2高變區內253位氨基酸或任何其他氨基酸的單一突變足以改變IBDV的毒力。在從接種了經典株疫苗的雞群中分離出來的vvIBDV中,觀察到G254D的甘氨酸-絲氨酸突變,因此該突變可能導致疫苗接種失敗。流行病學發現一個全新的獨特株,其在VP2高變區具有獨特的AA序列272T、289P及在VP1中234P位,這些序列是保守的,根據分子特征和致病性,將其稱為新型IBDV(dIBDV),廣泛分布在南美、歐洲、亞洲。
基于IBDV VP2分子流行病學的遺傳多樣性,學者建議采用新的分類法,將其分為7個基因型。一些基因型具有全球分布性,如基因1型(caIBDV)和基因3型(vvIBDV及其重組株)的成員,而基因2型(美洲發現的vaIBDV)、基因4型(南美洲發現的dIBDV)和基因5型(墨西哥發現,被認為是caIBDV和vaIBDV的重組株)的成員呈區域性分布。基因6型來自中東(沙特),與意大利的ITA基因型有92-93%的相關性,與俄羅斯的IBDV RF-5/94株有94-95%的相關性。基因7型的成員主要來自澳大利亞和少數俄羅斯。
三、血清型和致病型
IBDV在抗原上分為血清1型和血清2型。血清2型主要感染火雞,對雞無毒力,對血清1型也不能提供交叉保護。血清1型對雞具有不同致病性,這源于毒力、免疫抑制和抗原性的不同。早期的IBDV疫情主要是經典株(caIBDV)引起的,法氏囊炎癥、腫大,然后萎縮。20世紀80年代初,在美國、中美洲和澳大利亞出現變異株(vaIBDV),它在抗原上不同于經典株或超強毒株,只引起法氏囊萎縮,沒有炎癥。80年代末,西歐、東南亞和非洲出現的超強毒株(vvIBDV),特征是法氏囊腫大,然后萎縮,其毒力比經典株更強,死亡率超過70%。經典株和超強毒株會引起出血性炎癥,并伴有嚴重的法氏囊濾泡損耗,兩者僅在死亡率上有所不同(caIBDV為30-60%,vvIBDV為70-100%)。而變異株導致法氏囊快速萎縮,沒有炎癥、出血,死亡率10%以下或沒有。接種過caIBDV疫苗的雞群,仍然會感染變異株。變異株和超強株,以及最近新型IBDV(dIBDV)可以突破母源抗體,并感染青年雞,蛋雛雞死亡率高達60%,肉仔雞死亡率達25%。超強株雖然與經典株具有抗原相似性,但是經典株母源抗體很高的雞群,仍然會感染超強毒株。
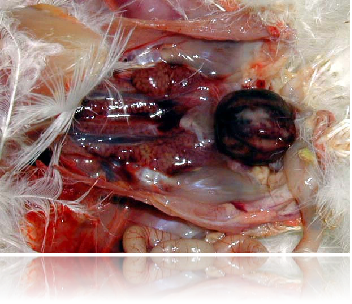
法氏囊出血
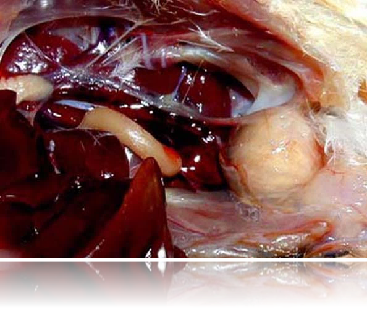
法氏囊腫大
目前,IBD疫苗主要是滅活苗或活疫苗。種雞接種了滅活苗或減毒活疫苗,雛雞通過母源抗體就獲得對IBD的保護。然而,母源抗體會干擾減毒活疫苗的有效性,除非使用中等毒力疫苗和強毒疫苗,這樣會造成疫苗引起的法氏囊損傷,導致免疫抑制。因此,迫切需要開發安全有效的疫苗,甚至在母源抗體存在的情況下,也能誘導正確的免疫應答。
四、先天免疫應答
實驗發現,使用變異株開發的減毒活疫苗和滅活疫苗,都對感染經典株或變異株的雞具有交叉保護作用;而使用經典IBDV株開發的疫苗部分保護或不能保護變異株的感染。
基因表達研究表明,急性感染IBDV,激活了法氏囊T細胞和脾臟巨噬細胞。法氏囊細胞被耗盡,與NK細胞、巨噬細胞和T細胞激活相關的基因上調,這些基因包括干擾素、白介素和MIP-1,以及調節先天免疫系統的基因,如MD-1、MD-2、補體、熱休克蛋白,以及炎性和促炎反應基因。
IBDV感染過程中先天免疫應答的遺傳調控
IBDV經口感染后的8-12小時,可在腸道的單核巨噬細胞中檢測到,然后這些細胞將病毒轉運到法氏囊,在帶IgM的B細胞內進行有效的病毒復制。感染激活NF-kB通路和其他細胞內信號通路。巨噬細胞大量浸潤法氏囊可誘導促炎介質如白介素的大量表達。在IBDV的研究中,觀察到中樞淋巴器官中IFN、趨化因子、補體成分、防御素的高表達。實驗研究發現,NK細胞可能參與了IBDV的感染發病和先天免疫應答。
李小慶
2023.03.08